In our earlier works 1 we have shown that an impulse through nerves is conducted electrically through orientations of water molecules, while ions present are responsible for maintaining the specific orientation. However, it remained unexplained as to how any sensory stimulus is received by a receptor before its transmission through the nerve.
It is a well known fact that there are five basic types of sensory receptors, identified on the basis of the kind of sensation they help in transmission. The most important feature of the sensory receptor, is that irrespective of the type of stimulus that excites it, it results in the alteration of electrical potential of membranes of the receptor, often termed as a receptor potential.
It is believed that receptors can be excited in one of several ways to cause receptor potentials:
They are usually classified 2 as :
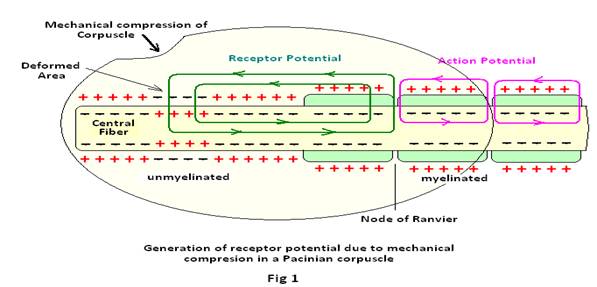
(1) mechanoreceptors, which detect mechanical compression or stretching of the receptor by mechanical deformation of the receptor that is believed to stretch the receptor membrane and open ion channels. The mechanism generating a receptor potential produced in the pacinian corpuscle from mechanoreceptors is depicted in Fig 1below. In the figure, the small area of the terminal fiber is deformed by mechanical compression of the corpuscle. The Na+ ions are allowed to diffuse to the interior of the fiber through opening of the ion channels. The diffusion of positive ions increases positivity inside the fiber and generates receptor potential. A local circuit of current flow is then induced by the receptor potential (shown by arrows) that propagates further along the nerve fiber. The local current flow then depolarizes the fiber membrane at the first node of Ranvier, that itself lies inside the capsule of the pacinian corpuscle. The depolarization of the fiber membrane then initiates the action potential for further propagation along the nerve fiber.
(2) thermoreceptors, which detect variation in temperature ( some receptors detecting cold while others warmth ) by alteration in the permeability of the membrane. It is believed that the cold and warmth receptors are stimulated by changes in their metabolic rates, and that these changes result from the fact that temperature alters the rate of intracellular chemical reactions more than two fold for each 10°C change. Thus in thermo receptors it is not the direct physical effects of heat or cold but the chemical changes due to temperature variations that stimulate the nerve endings.
(3) nociceptors (pain receptors), which detect damage (physical/chemical) occurring in the tissues by mechanical deformation of the receptor or application of a chemical to the membrane.
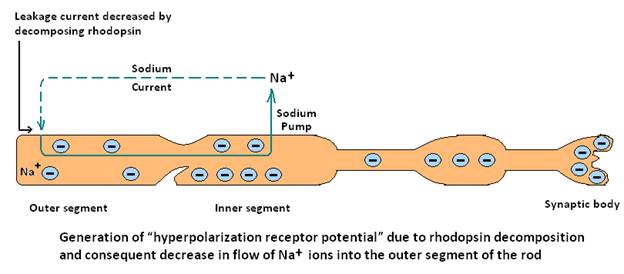
(4) electromagnetic receptors, which detect light on the retina of the eye, by the effects of electromagnetic radiation, such as light on a retinal visual receptor, which either directly or indirectly changes the receptor membrane characteristics and allows ions to flow through membrane channels. In the eye, the receptor potential generated on exposure of the rod to light is quite different from receptor potential generated in other sensory receptors (i.e. depolarization). It is believed that excitation of the rod due to light causes increased negativity of the intra rod membrane potential which is a state of hyperpolarization, meaning more negativity than normal inside the rod membrane.Thus this process of increased negativity inside the fiber is exactly opposite to the decreased negativity (i.e. depolarization) that is found in other sensory receptors. Actually the decomposition of rhodopsin decreases the rod membrane conductance for Na+ ions in the outer segment of the rod to the interior of the rod, though Na+ ions are continually pumped outward through the membrane of the inner segment .This leaves more Na+ ions (i.e. positive charge) in the rod than leaking back in to create increased negativity inside the membrane (i.e. hyperpolarization).
(5) chemoreceptors, which detect taste in the mouth, smell in the nose etc. by application of a chemical to the membrane, which also opens ion channels.
The above theory of the generation of receptor potential and conduction of electrical impulses to the brain through nerves cannot however explain:
i) any of the hallucinations i.e visual, auditory, olfactory, gustatory, thermoceptive , nociceptive or tactile, where a specific sense is perceived in the absence of any evident stimulus. For example, in olfactory hallucination the odor of rotten eggs, urine, vomit etc are often perceived in the actual absence of such substance.
ii) conduction of visual impulses in the eye where the Rod Receptor Potential is believed to be Hyperpolarizing (i.e. inside of the fiber is more negative than normally in a nerve fiber) but not Depolarizing.How such excess negative charges become stabilized inside the fiber remains a question. Moreover, in generation of other receptor potentials, the positive electrical charges carried by the inward-diffusing sodium ions are believed to help formation of initial depolarized area.Due to dissimilar polarity, a local current is believed to flow and a local circuit (shown by arrows) is formed (Fig 1). Therefore, induced opening of the sodium channels in these new regions helps the rapid spread of receptor potential in that region.The receptor potential in turn induces a local circuit of current flow that spreads along the nerve fiber through the similar mechanism.
Thus it is clear that generation of receptor potential is also initiated by inward diffusion of Na+ions as in action potential for propagation of impulses1. But it is not clear in case of the eye, after generation of receptor potential through hyperpolarization Fig 2 (i.e when the rhodopsin in the outer segment of the rod is exposed to light there is marked negativity inside due to regular outside diffusion of Na+ions at the inner segment while restricted inward diffusion at outer segment) how such potential initiates action potential in nerve fibers for further propagation of the electrical impulse as found for other receptor potentials (Fig1.)
iii) how any electrical impulse generates receptor potential at the site of the receptor and is conveyed through the nerves. Obviously, though the positive part of the impulse ( i.e. positive electrode) would repel Na+ ions and help opening of the ion channel, on the other end the negative part of the impulse (i.e negative electrode) would attract the Na+ ion and hence would restrict opening of ion channel and consequentlyany electrical conduction1.
iv) conduction of alternating current (A.C.) through body as the polarity of the electrode connected to the A.C. source reverses with time. Thus for Conduction of A.C. through body3 it appears that conduction by nerves occurs through both depolarization & hyperpolarization !
v) conduction of radio impulse through the body seems very interesting as here radio waves ( electromagnetic radiations) are carried through the body to the receiver of a radio, the body behaving as an antenna. Antenna are made of metals and EMR induces a flow of electrons hence emf and a small alternating current. The electric field & magnetic field are oscillating in phase perpendicular to each other and A. C. is produced . So when the body acts as an antenna it must be transforming the EMR into small alternating currents as done by an actual metallic antenna. Now it is a question how the body produces and conducts such induced alternating current, in the presence of the EMR, when it acts as an antenna for radio. It appears that such induced A.C. must be therefore carried through nerves by both depolarization & hyperpolarization !4
The mechanism of both the generation of receptor potential and initiation and subsequent propagation of action potential is depolarization in the case of a pacinian cell. However, it greatly differs for the eye, wherein it is stated to be hyperpolarization at receptor sites. Therefore the question arises how the electrical impulse is further carried to the brain through the nerves that are known to carry impulses through depolarization ?
To show that change in orientations of water molecules at one side of
the semi permeable membrane certainly influences the orientations of water
molecules on the other side of the membrane, the following experiment is
performed:
Experiment:
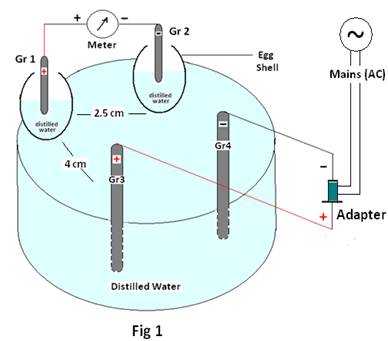
Condition1:Two graphite electrodes Gr1 & Gr 2 are placed in two different egg shells (about 2.5cm apart) containing distilled water(10 ml) and are placed in a glass bowl containing about 60 ml of distilled water. The emf generated by the Graphite pair (Gr1 & Gr 2) is measured and recorded as initial emf. The current from an external source (Adaptor) is allowed to pass through two different electrodes Gr3 & Gr4 placed in the glass bowl (as shown in Fig 1) at a distance of about 4 to 4.5 cm from the other pair. Now the emf of the Gr1 & Gr2 is recorded as final emf for each applied emf of the adaptor and recorded in table1 below.
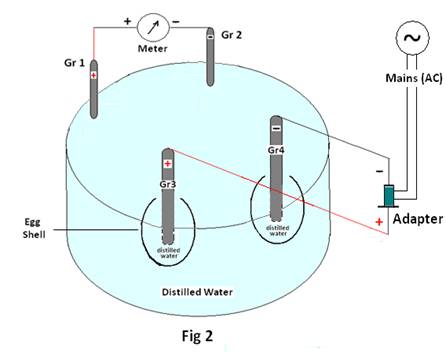
Condition2:Two graphite electrodes Gr1 & Gr 2 are placed in the glass bowl containing about 60ml of distilled water (i.e. without any egg shell). The emf generated by the Graphite pair (Gr1 & Gr 2) is measured and recorded as initial emf. The current from an external source (Adaptor) is allowed to pass through two different electrodes Gr3 & Gr4 placed in two different egg shells containing distilled water (10ml) and placed in a glass bowl (as shown in Fig 2).Now the emf of the Gr1 & Gr2 is recorded as final emf for each applied emf of the adaptor and recorded in table1 below.
| Pointer on Adapter showing emf (in volts) | Actual emf shown at Gr3-Gr4(in volts) | Condition 1 | Condition 2 | |||
| Gr1 & Gr2 are placed in Egg ShellWhile Gr3 & Gr4 directly in the bowl containing distilled water | Gr1 & Gr2 are placed directly in bowl containing distilled water While Gr3 & Gr4 in the Egg Shells | |||||
| Initial emf of Gr1-Gr2 21 mV | Initial emf of Gr1-Gr2 60 mV | |||||
| Final emfGr1-Gr2 (in mv) | Difference(in mv) | Final emfGr1-Gr2 (in mv) | Difference(in mv) | |||
| 1 | 1.5 | 2.02 | 80 | 59 | 69 | 9 |
| 2 | 3.0 | 4.46 | 186 | 165 | 78 | 18 |
| 3 | 4.5 | 6.92 | 280 | 259 | 86 | 26 |
| 4 | 6.0 | 8.91 | 370 | 349 | 95 | 35 |
| 5 | 7.5 | 10.73 | 450 | 429 | 103 | 43 |
| 6 | 9.0 | 12.44 | 550 | 529 | 114 | 54 |
| 7 | 12.0 | 14.81 | 669 | 648 | 123 | 63 |
Observations: On passing of current from external source through the graphite electrodes Gr3 & Gr4, the final emf of the electrodes Gr1 & Gr2 is found to increase for both the applied condition 1 and condition 2.
Discussion: Though final emf recorded for a given applied emf (i.e. from external source) varies for condition 1 & condition 2, it is fact that in spite of electrodes being placed in egg shell (Gr1, Gr2 and Gr3, Gr4 for condition 1 & condition 2 respectively) containing semi permeable membrane, the change in emf is very considerable. Now a graph is plotted for the emf generated at Gr1 & Gr2 against the applied emf at Gr3 & Gr4.
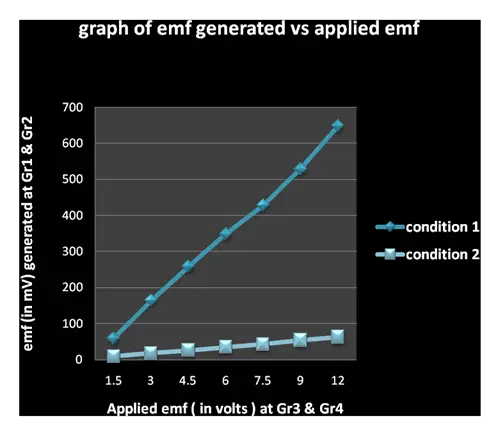
The varying emf recorded at Gr1& Gr2 in condition 1 & 2 for various applied emf at Gr3 & Gr 4 shows that the former varies directly to the latter. The comparative steep slope found in condition 1 than in condition 2 indicates that the presence of a semi permeable membrane plays a key role in both the maintenance and propagation of induced orientations.
Moreover, it is now well known that any generation of emf by a pair of electrodes in water is due to alteration in orientations of water molecules5. Similarly, generation of any particular emf at Gr1 & Gr2, within the eggshell owing to application of particular emf from external source for condition 1 and outside the eggshell owing to application of particular emf for condition2, can easily be attributed to the specific change in orientations of water molecules.
From above it is clear that any change in orientations of water molecules in the vicinity of the electrodes kept within the eggshells containing semi-permeable membrane can definitely bring a change in orientations of water molecules within the egg shell.
We have to consider those factors that are able to bring change in orientations of water molecules in the vicinity of the electrodes kept within the eggshell and hence are able to generate the receptor potential:
# application of chemicals in water can bring change in orientations of water molecules 6. This property of chemicals to bring change in orientations of water molecules help explain generation of receptor potential (electrical impulse) to detect taste in the mouth, smell in the nose etc. through chemical applications. The chemical changes as a result of variations in temperature can also bring change in orientations of water molecules and thus help generation of receptor potential in case of thermo receptors. In the case of the eye, there is chemical change, due to decomposition of Rhodopsin, that brings change in orientations of water molecules for generation of receptor potential.
# application of electrical energy in water can also bring change in orientations of water molecules 6,7. Thus any sort of electrical imbalance in water can bring change in its orientations. This property is seen in case of mechanical deformation where due to the diffusion of positive ions inside the fiber, positivity increases inside the fiber. As a result there is an electrical imbalance and consequent change in orientations of water molecules in the vicinity of receptors and thus helps generation of receptor potential in case of mechano receptors.
Thus the alteration in emf on application of heat, mechanical deformation (touch), addition of salt (chemical) and passage of electrical energy, confirm their ability to bring changes in orientations of water molecules at the site of the sensory receptor for acting as electrical impulse.
Inference:
The change in orientations of water molecules, brought in the vicinity of a sensory receptor by the sensory impulse, generates the receptor potential for transmission of sensory impulse.
References
[1] Conduction of Electrical Impulses Through Nerves – An Explanation,Ruhul Amin & Biplab Chakraborty (Published in Homeopathy for Everyone Apr2014 hpathy.com
[2] Textbook of Medical Physiology (Eleventh Edition) -Guyton and Hall
[3] The Phenomenon of Electrical Earthing Explained By Orientations of Water Molecules, Ruhul Amin & Biplab Chakraborty (Published in Homeopathy for Everyone Jan2014
[4] The Electromagnetic Radiation Induces Electric Current in Living body by delocalization of electrons through water molecules, Ruhul Amin & Biplab Chakraborty(Published in Homeopathy for Everyone June 2014hpathy.com
[5] The Driving Force Behind Homeopathy – Explaining the Principle Behind the Generation of Emf in Water,Ruhul Amin & Biplab Chakraborty (Published in Homeopathy for Everyone Nov2013 hpathy.com
[6] Solutions Ionic or Non-Ionic Conduct & Retain Electrical Energy,Ruhul Amin & Biplab Chakraborty Homeopathy for Everyone July2012 hpathy.com
[7] The Key to Homeopathic Dilution, Ruhul Amin & Biplab Chakraborty (Published in Homeopathy for Everyone Nov2012 hpathy.com
[8] Wikipedia



This is a very interesting document and I kindly salute it.
Now, the point is that the signal goes to the brain and that is the healing effect of homeopathy, something I say in my books. (Sorry, only in Spanish. 🙁 )
Sir,
Thanks. We request you to incorporate our findings in your publications , if possible, so that your readers and Spanish speaking population at large can be made acquainted with it.
Ruhul & Biplab