1. Introduction
Previous work (Hart, 1985) showed that in vivo dielectric measurements on plant tissues can give stable readings which indicate the health of the plant and even show variations following injury and subsequent healing. The present paper describes a simple apparatus to monitor variations in the electrical resistance of plants. The results of preliminary measurements are correlated with the lighting and electromagnetic environment.
Two plant species chosen were representative of two very different growing environments. : Crassula argenta (Money of Jade) is a hardy succulent preferring warm light and dry conditions. The leaves are moist and approximately 700 sq. mm. in area. The other, Dieffenbachia picta superba (Leopard Lilly) is a tropical plant and enjoys shady moist and warm conditions. The leaves are 1mm thick and 500 – 1500 sq. mm. in area. The plants were purchased from a local supermarket.
2. Apparatus
The basic problems in the design of apparatus to measure in vivo the electrical properties of plant tissues are firstly, the wide range of external factors which may affect the physiological state of the plant system and secondly, the possibility that the apparatus may contribute to these particularly through the variation in the electrode characteristics and physiological effects due to small currents entering the plant through the electrodes.
Preliminary experiments were made using electrodes in contact with the leaves and inserted into the stem. Although inserting an electrode into the stem produced localised damage, the wound soon healed and thereafter gave a stable contact if a chlorided-silver wire was used. With electrodes applied to the leaf, electrode noise, effects of mechanical disturbance and tissue damage produced by electrode jelly combined to give unsatisfactory results. Accordingly, an electrode of chlorided-sliver wire 2 mm in diameter was inserted into the stem of each plant to be measured at about 20 mm above the soil level (Fensom, 1963; Carter and Blanchard, 1978). These were left in position for several days for the wound to heal.
The other electrode was a 2 mm. diameter wire inserted into the plant pot to a depth of about 50 mm. This was of copper which had been ‘tinned’ with a tin-lead solder. To reduce electrical interference, the plants were connected to the measuring circuit by a coaxial cable. The inner conductor was connected by a clip to the stem electrode. The outer conductor (brading) was connected to the soil electrode.
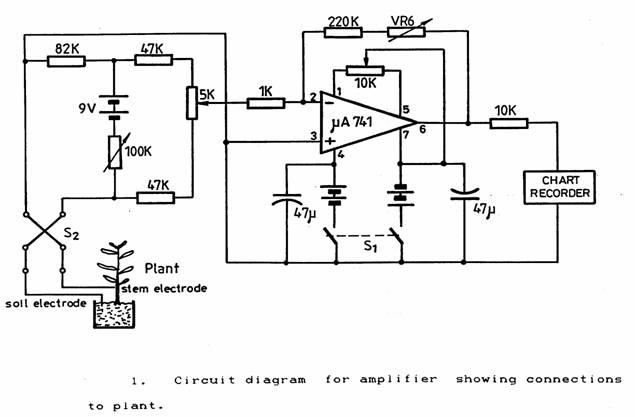
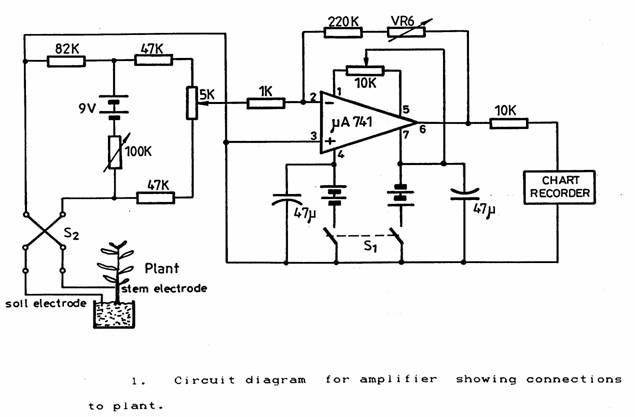
Initial tests indicated that the resistance from the stem electrode to the soil electrode would be between 40 k? and 100 k?. The circuit used is shown in Figure 1. The switch S2 is a polarity reversing switch that permits the periodic polarity reversal of the 50 µA of bridge current flowing through the plant from the bridge circuit. This level of current appeared to be satisfactory although some of the stems on the Dieffenbachia produced more shoots when a current was flowing. The 100 k? variable resistance can be set to adjust the current in the plant according to its resistance. The 5 k? potentiometer gives fine adjustment of the bridge null-point. The amplifier was an inexpensive µ741 operational amplifier. The bias conditions are a compromise between the limitations of the amplifier off-set current and the requirement to minimise the current passing through the plant stem. The gain of the amplifier could be adjusted by adding the variable resistor VR6 to the 200 k?. The bridge and amplifier were independent of the mains supply being run from the three 9V batteries shown. The output was connected to a chopper-bar chart recorder (Goerz-Electro Model 226253) running on the 5 mA range at a speed of 20 mm/hr and sampling at 3 second intervals.
Stability was tested by connecting a 40 k? resistor to the input cable instead of the plant. No variation or drift was visible on the chart over an 8-hour period. The bridge circuit used is not linear when operated away from balance conditions. For plant resistances between 40 k? and 80 k? the maximum non-linearity on the chart amounted to an acceptable ± 6%.
3. Tests on the Plants
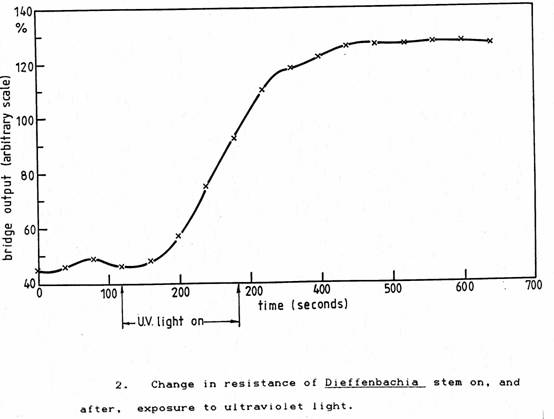
To test the overall reaction rate of the plant and measurement system, a specimen of Dieffenbachia in which a stem electrode had been inserted and stabilised was connected to the bridge input cable. When the bridge output had become steady (the electrode time constant was about 10 seconds) the plant was exposed to ultra-violet light from a fluorescent tube for 2 minutes from a distance of ½ metre. There was a latency period of 1 minute after which the change in bridge output had a time constant of about 3½ minutes as shown in Figure 2. The decay of the response was much slower.
The two plant species used were placed day and night on a window ledge in the laboratory as far as possible from electrical apparatus and where they would receive the normal environmental fluctuations of light and temperature. Their stem resistances were found to vary in synchronism as shown in Figures 3a and 3b.
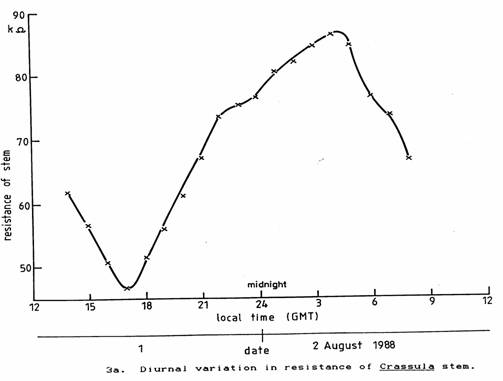
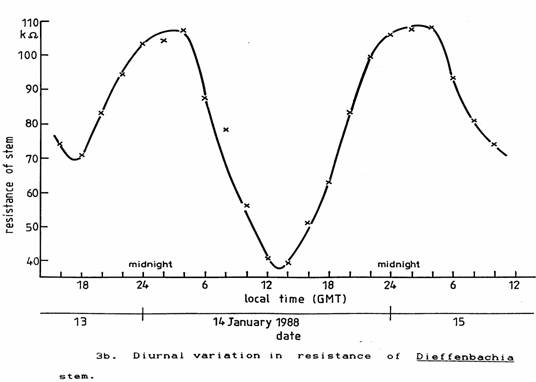
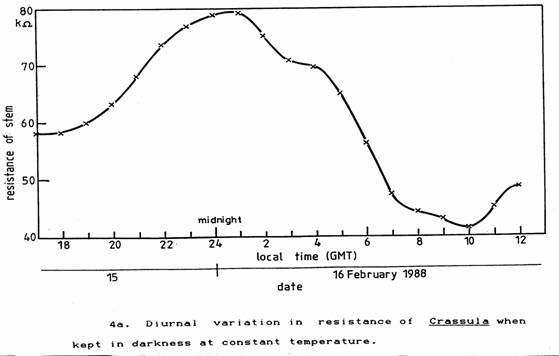
To eliminate the variations of light and temperature the plants were placed inside an incubator set to 18 ± 0.1 °C giving shielding from light and all other electric and electromagnetic fields except for ELF and steady magnetic fields. The observed periodic variation in the resistance persisted as seen in Figures 4a and 4b. Although the plants started to prepare for night as soon as they had been placed in the incubator, they both commenced to prepare for daylight and photosynthesis between 3 and 4 am (GMT), that is about 2 hours before local twilight (Reed, 1988). In England, in February the twilight extends about half-an-hour either side of sunrise and sunset.

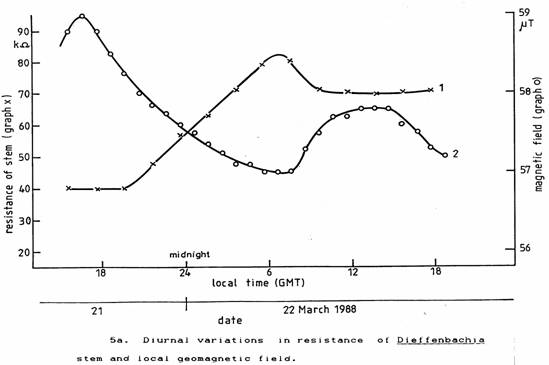
Since only ELF and magnetic field fluctuations could have reached the plant within the incubator to act as a ‘zeitgeber’, the plants were again placed on the window ledge in normal light and indoor temperature conditions. Then, the local geomagnetic field was measured simultaneously with the plant stem resistance. There was a close correlation between these two parameters as seen in Figures 5a and 5b. The plant stem resistance changes lagged about 1½ hours behind the geomagnetic field changes.
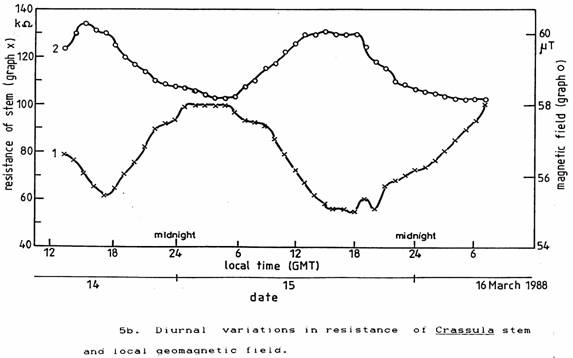
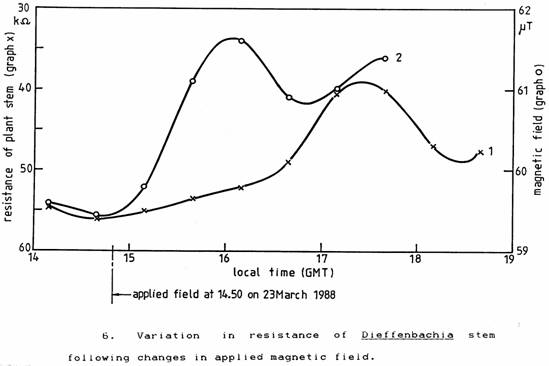
To confirm that a changing magnetic field could produce such an effect, the plants were placed in the magnetic field generated by a steady current in a large coil arranged so as to simulate the measured changes in the geomagnetic field. The results shown in Figure 6 confirmed that the observed plant stem resistance changes could be produced by a changing magnetic field and again gave a time lag of 1½ hours.
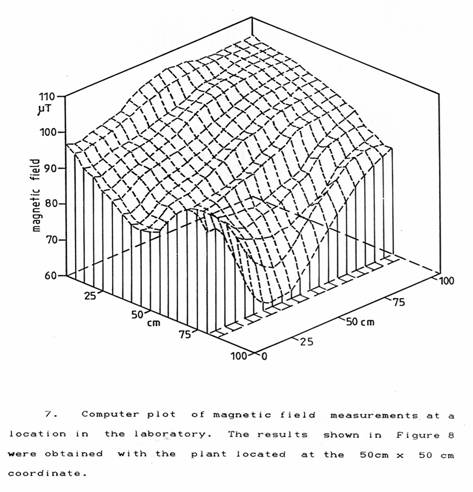
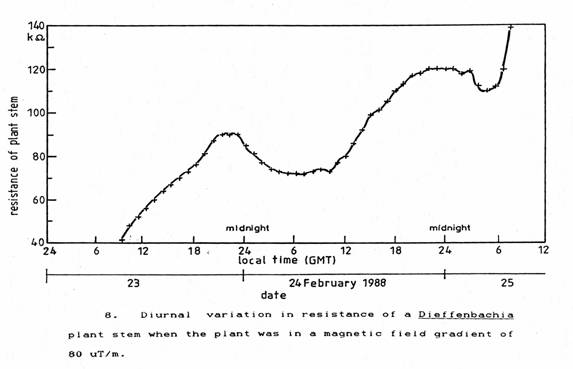
To try to suppress the effect of this ‘zeitgeber’ a Dieffenbachia plant was placed on the floor in a part of the laboratory close to a steel floor joist where there was a strong gradient of magnetic field. This was measured using a geo-magnetometer with a graphics computer (BPM2001, Bio-Physik Mersmann ). The resulting plot shown in Figure 7 gives a magnetic field gradient of 80 µT/m. which represents a variation of 3 nT across a leaf of the plant and 1 nT across its stem diameter. Since the plants were responding to changes in the geomagnetic field of total magnitude 1 µT and began to respond at changes of 100 nT it is not so remarkable that this gradient of magnetic field was able to paralyse the ability of the plant to photosynthesise properly. The plant was placed at the 50 cm × 50 cm coordinate shown in Figure 7. At this location, the overall resistance of the stem as shown in Figure 8 increased steadily over 48 hours. The periods of decreasing resistance occurred much later in each succeeding day and for shorter durations. At the end of 48 hours, the plant had wilted, the leaves had become discoloured and overall the plant appeared to be in very poor condition. Subsequently, it recovered on being moved back to its former location on the laboratory window ledge.
4. Conclusions
From the above preliminary experiments on two plant species preferring very different habitats, it appears that the electrical resistance between an electrode inserted into the stem and one inserted into the potting medium is a good indicator of the plants’ activity and impending activity. The resistance changes for the plant indoors can be stimulated rapidly by exposure to ultra-violet light. Diurnal variations in the stem resistance persist when the plant is kept inside an incubator in darkness and screened from all but ELF and geomagnetic fields. These variations correlate with the diurnal variations in the local geomagnetic field, but they lag in phase by about 1½ hours. Comparable variations in stem resistance can be stimulated by equivalent laboratory generated magnetic field changes and rates of change, both have to be correct. Finally, the effect of exposure to a magnetic field gradient giving a magnetic field variation over the dimensions of the plant which is greater that the geomagnetic fluctuations providing its ‘zeitgeber,’ results in a rapid decline in its photosynthesis activity and general vitality.
Acknowledgments
Thanks are due to Dr. Ludger Mersmann for the loan of the geo-magnetometer and computer and to Dr. Kaydar Quboa for making the magnetic field plot which forms Figure 7.
References
Carter JK and Blanchard RO (1978) Electrical resistance related to phloem width in red maple. Canad. J. For. Res. 8: 90-93.
Fensom DS (1963) The bioelectric potentials of plants and their functional significance. Canad. J. Botany 41: 831-851.
Hart F-X (1985) The extremely low frequency electrical properties of plant stems. Bioelectromagnetics 6: 243-256.
Reed’s 57th. Nautical Almanac (1988) Thomas Reed: New Malden.
Appendix 2.
Frequency, Coherence and Response Rate Theory
Anything in a state of oscillation has a variation which usually repeats in cycles according to the mathematical relation called a sine function, hence the term sine-wave. This function is the mathematical solution of an equation describing an oscillation. If the ‘sine’ had not previously been found in trigonometry it would have had to be invented for this equation.
The number of cycles per second is the frequency of the oscillation. Equations tend to give frequencies in radians per second and since there are 2? radians in one cycle or circle this factor often appears. Two waves can have the same frequency and wavelength but they may still have a difference in phase. If the phases of two waves of the same frequency differed by half a cycle they would be mirror images and if combined they would cancel. This is called destructive interference. Coherence can be regarded as a measure of the capacity of two waves to interfere.
If the effect of an oscillation is propagating through space, the distance travelled per cycle is the wavelength, and frequency multiplied by wavelength gives its velocity of propagation.
The band-width of a resonance is usually measured between the half-power (3db) points. The more precise the frequency (i.e. the narrower the band-width) the longer the resonance takes to build-up and to decay (echo). This is often described and measured by a parameter called ‘Q‘ (= frequency / bandwidth). These are all mathematically related as shown in Figure 1.
The early part of the 20th. Century saw Planck’s hypothesis that oscillators can only emit or absorb energy in quanta and not in any arbitrary amount as assumed in classical physics. Bohr connected these oscillators with atomic structures, thereby explaining why atoms do not ‘run down’. Hahnemann’s original potencies did not ‘run down’ and they were found to be clinically effective 150 years after he had prepared them. Homeopathic potencies must involve some macroscopic type of quantum physics to provide a similar stability to that given by quantum physics at the atomic level. The Classical Electromagnetic Field describes physical states for which the phase is well defined, but the number of particles (quanta) is undefined. It can describe the propagation of electromagnetic radiation but, as Einstein postulated, quanta are inherent in the nature of the radiation itself. Froehlich pioneered the concept of coherence in living systems. Del Giudice and Preparata showed that domains of coherence were a fundamental property of water.
There is some very interesting physics behind homeopathy. It involves the interaction with living systems of frequencies stored in coherence domains in a homeopathic potency.
Figure 1
Resonance Build-Up and Decay in Classical Physics
The quality (Q) of a resonance expresses the sharpness of a resonance and the response rate. It is the resonant frequency divided by the bandwidth at the half-power (3dB) points and is the ratio: (energy stored / energy lost per cycle).
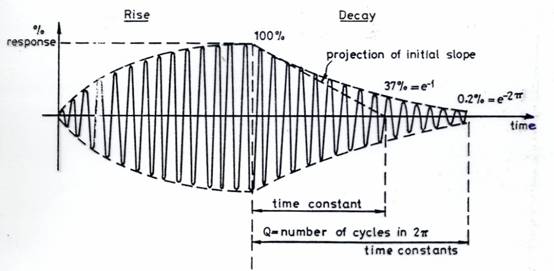
Within a coherence domain, the coherence in the phase increases as more particles in the domain are allowed to fluctuate randomly. This takes up the Heisenberg Uncertainty. It is this uncertainty which limits how narrow the bandwidth can be.
The bandwidth of a frequency can be measured with sufficient precision. This gives the number of particles involved in fluctuation which in turn gives the total number of particles within the coherence domain.
If these particles are protons they will spin and the spin axis will precess in a magnetic field. This precession is a current loop which will generate a local magnetic field. There is a certain number of protons which must precess coherently to satisfy the proton nuclear magnetic resonance conditions for any frequency of precession. In other words, if the protons are started precessing at any given frequency, they will generate exactly that magnetic field which will keep them precessing at that frequency.
This memory of frequency will be stable unless the entire coherence is broken up by removing the geomagnetic field. This erases a homeopathic potency. The critical magnetic field at which this happens gives the size of a coherence domain as 53µm. This in turn gives the number of protons available to remember a frequency. There are enough protons in a domain to remember over 800 different frequencies which is what is measured by multiple imprinting. The numbers derived from experiment and theory all fit.
Within a coherent system, external radiation will interact with an entire coherence domain or, not interact at all. It is the interaction and scattering by individual molecules which gives matter its refractive index. If radiation does not interact, it travels with the free-space velocity of light. If it does interact, it will interact with an ensemble of molecules oscillating in-phase as a massive coherence domain. This greatly reduces the velocity and is equivalent to an enormous refractive index.
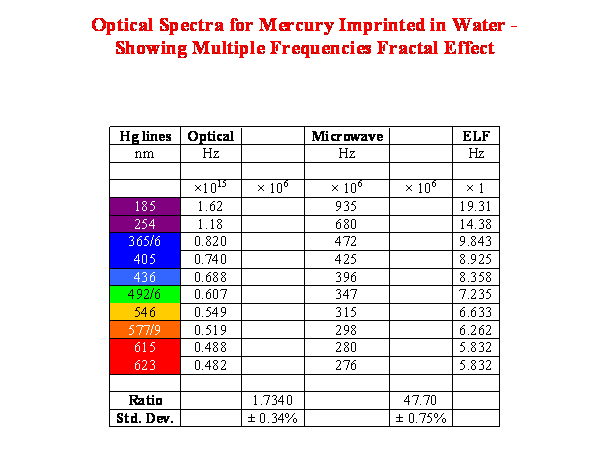
Fractality arises from frequencies within a coherence domain. The constant parameter becomes the coherence length and this makes frequency proportional to any velocity with which the coherence can propagate. Table 1 shows the fractal frequencies generated by imprinting the optical spectrum from a mercury discharge lamp into water. Combining the frequency ratios gives the ratio: Optical/ELF = 8.25×1013.
If a plant wants to do something quickly or needing much energy, it can use the ultra-violet. If it wants to ignore irrelevant environmental fluctuations it can chose a lower fractal of frequency, yet still retain the capability to switch over to a fast activity or a high energy activity like photochemistry.
Table 1
General References
Arani, R. Bono, I. Del Giudice, E. Preparata, G. (1995) QED Coherence and the Thermodynamics of Water. Intl. J. of Mod. Phys. B, 9, 1813-1841.
Froehlich, H. (1983) Coherence in Biology, in ‘Coherent Excitations in Biological Systems’, Froehlich, H. and Kremer, F. (Editors). Berlin: Springer-Verlag pp 1-5.
Froehlich, H. (1988) Theoretical Physics and Biology, in Froehlich, H. (Editor) “Biological Coherence and Response to External Stimuli”. Berlin: Springer-Verlag pp 1-24.
Preparata G. QED Coherence in Matter. Singapore: World Scientific, 1995.
Smith CW. (1998) Is a living system a macroscopic quantum system? Frontier Perspectives, 7(1), 9-15, (Temple University, Philadelphia, 1997 Lecture at Frontier Sciences Department).
Smith CW. (2007) Water – its clinical and scientific depths. In: Emoto M, The Healing Power of Water. London: Hay House. Chap.3, pp 77-88.
Smith CW (2008) Froehlich’s Interpretation of Biology through Theoretical Physics. In: Hyland GJ and Rowlands P (Eds.) Herbert Froehlich FRS: A physicist ahead of his time. Liverpool: University of Liverpool, 2nd edition, pp 107-154.
[1] Gurwitsch AG and Gurwitsch LD “Twenty Years of Mitogenetic Radiation: Emergence, Development and Perspectives” 21st. Century, Fall 1999, pp.41-53 also appended to Proc. Intl. AG Gurwitsch Conf. Sept 28- Oct2, 1994 as English translations of a 1943 article in Russian.
[2] hpathy.com “Homeopathy – How it works and how it is done” January to July 2008.
[3] Recommendations mostly taken from “Agro-Homeopathy – An Alternative for Agriculture” by Dr. Niurka Meneses Moreno in hpathy.com December 2008 and reproduced with permission.
[4] hpathy.com “Homeopathy – How it works and how it is done”. Chapter 5, Section.8, May 2008.
————————————————-
Cyril Smith was born in London, England, in 1930. He started work in radar in 1947; he was a Research Fellow at Imperial College, London, from 1956 under Blackett and McGee working on Medical X-ray Images. From 1964 at Salford University in the Electrical Engineering Department, his research activity included: Instrument Technology, Medical Electronics, Dielectric Liquids, Electromagnetic Effects in Living Systems and Water. In 1973, his co-operation with Herbert Froehlich, FRS commenced. In 1982, he first became involved with the diagnosis and treatment of electromagnetically hypersensitive patients. He was Secretary of The Dielectrics Society from 1972-1983. In 1989/90, his co-authored book “Electromagnetic Man” was published. That year he also took early retirement. He continues to be active in research and writing. For a bibliography of his writing, see chapter one.